

植物光合系统既要在多云或阴天低光照条件下保持高效捕光和传能效率,又要在正午强光持续照射下避免由此引发的氧化损伤即光保护。植物的光保护功能是将过剩的激发能以热的形式耗散掉。 在自然环境中,太阳光的辐照强度可以在短时间内呈现出十几倍的涨落。当云的阴影遮蔽住受强光辐照的叶片后,叶片还会将光保护状态持续数分钟之久,然后才切换到捕光状态。理论计算表明,由于状态开关切换的滞后效应,可以导致20%的光能损失。2016年,美国科学家通过遗传基因工程,获得开关恢复速率加快的突变株,发现其净光合效率比野生型增加约15% (Science, 2016,354,857)。
那么光合系统是如何调控蛋白质分子空间结构以快速响应环境光照条件的变化,实现低光照条件下高效能量传递及接近90%的电荷分离量子效率、并在强光照条件下快速切断传能通道进入光保护状态的?这个问题困扰了科学家近半个世纪,对该问题的回答并阐明其微观机理对于分子育种以提高农作物产量具有重大的指导意义。
中国科学院物理研究所/北京凝聚态物理国家研究中心软物质物理实验室翁羽翔课题组,深圳湾实验室量子生物学、美国明尼苏达大学化学系高加力课题组和Gianlugi Veglia教授合作,应用超快时间分辨荧光光谱及脉冲升温(T-jump)-纳秒时间分辨中红外瞬态光谱结合全原子分子动力学模拟及量子化学计算,实验与理论密切配合,揭示了高等植物光系统II主要捕光天线蛋白色素复合物(Light Harvesting Complex II,LHCII)三聚体,作为蛋白质分子机器,是如何实现高效捕光和光保护功能间可逆切换的机理。该研究表明,强光照射下的局部升温和光解水导致的光合膜囊腔侧酸度的增加所产生的驱动力,在微秒时间尺度内诱导LHCII蛋白质变构运动,导致捕光色素对叶黄素-叶绿素间的距离减小,发生叶绿素激发态被叶黄素分子淬灭的能量转移过程,由此将多余的激发态能量有效地转换成热能而耗散掉,从而实现了由高效捕光到激发态能量耗散状态间的快速切换。这一分子机器的作用机理密切关联了光合系统外部环境条件(环境温度,水的蒸腾作用,光合膜的硬度)、内部分子层次动态结构变化、以及植物表观现象的因果关系,解决了光合体系如何可逆地实现由低光照条件下的高效捕光/传能功能切换到高光照条件下的光保护功能这一个核心科学问题。
该研究揭示,聚集态的LHCII三聚体的荧光寿命和强度受温度和酸度调控,温度升高或者酸度增加都能够导致显著的荧光淬灭现象。通过荧光寿命分析表明,LHCII存在两种构像体,即长寿命的捕光构像(light-harvesting, H)和短寿命的能量淬灭构像体(Quenching, Q)。另外系统在较高的温度下通常还存在变性的蛋白构像(Denatured, D)。因而在变温过程中,系统中存在三种构像体,即H、Q 和D。结合荧光寿命和强度分析,可以严格得到三者相对含量随温度的变化关系。结果表明,淬灭态Q的含量随温度升高而增加,在55℃左右,达到最大值 (图1)。另外,蛋白的热稳定性分析表明,LHCII三聚体在55℃以下的结构变化几乎是完全可逆的。
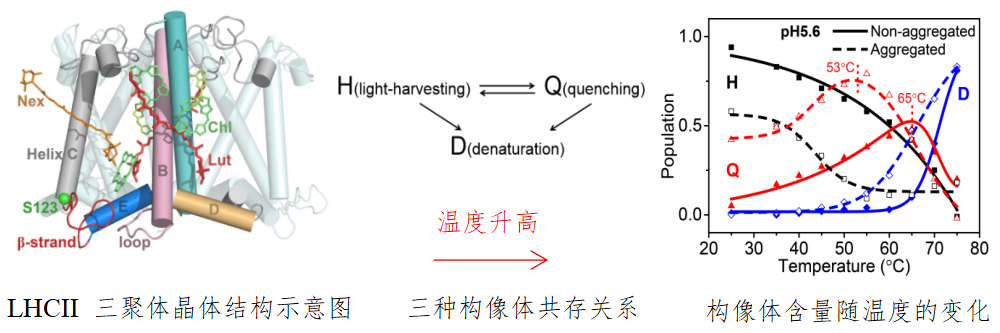
图1. 升温促使LHCII三聚体由传能状态到激发能淬灭状态的转换示意图。
随后应用自主研发的高精度脉冲升温—纳秒时间分辨中红外光谱仪(Rev. Sci. Instr. 2015,86,053105),结合变温傅里叶变换红外光谱,研究了LHCII三聚体在热胁迫下发生的结构变化,指认了囊腔侧处于部分亲水的310螺旋E(Helix E)的二级结构谱峰位置,并通过单点突变重组蛋白(S123G)进一步证实了对关键二级结构310螺旋结构E的光谱指认。实验表明温度升高与酸度增加都会促使囊体腔侧一段处于松弛状态的310螺旋E向结构更紧凑的α-螺旋转变。热胁迫下310螺旋E向α-螺旋转变的时间约1微秒,新生的α-螺旋向疏水内核区插入的时间约1.24微秒 (图2)。
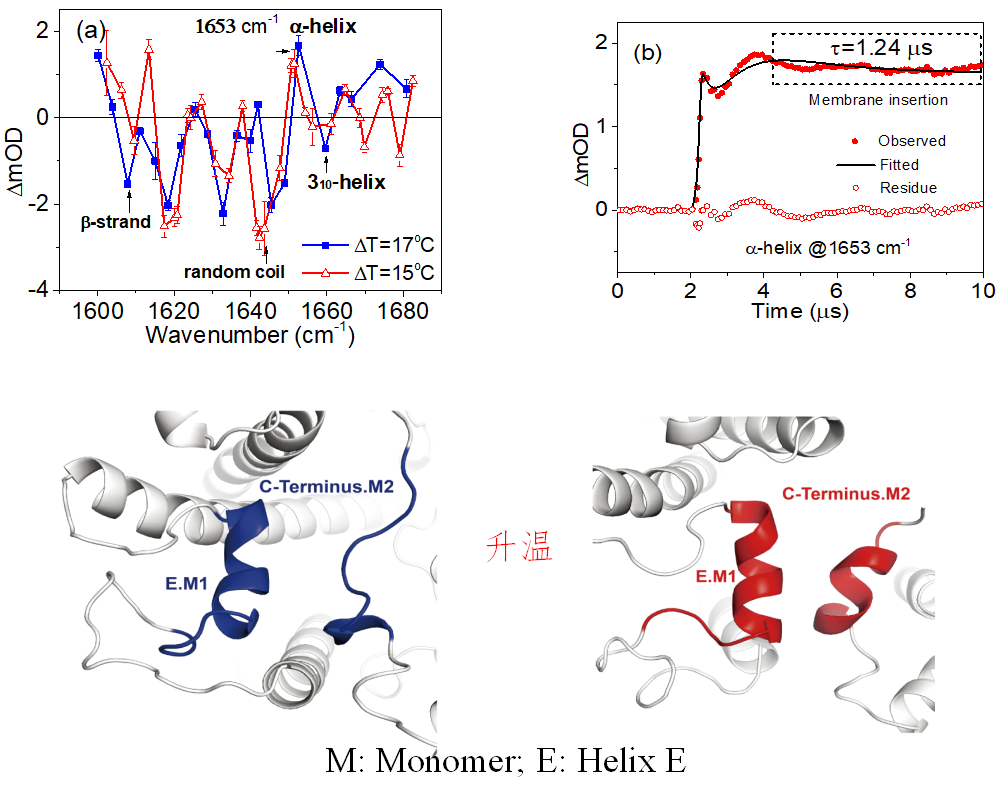
图2. 脉冲升温—时间分辨中红外光谱及动力学(a,b)(上半图)。实验表明新生α-螺旋插膜过程的时间尺度为1.24 μs (b)。分子动力学模拟结果(下半图)。分子动力学模拟揭示升温过程中处于松弛状态的310螺旋结构(E.M1: Helix E in Mononomer 1,蓝色部分)和相邻单体中的一段无规卷曲结构(C-terminus in Mononomer 2,蓝色部分)会协同转变为α-螺旋(红色部分)。
全原子分子动力学模拟结果表明,在升温过程中,蛋白质部分结构单元会发生失水作用,导致结构单元间的疏水作用增强,引起310螺旋E和相邻的LHCII单体中的一段无规卷曲结构协同转变为α-螺旋,并引起α-螺旋E和D向疏水核内部插入运动,从而在原子尺度上阐释了脉冲升温—时间分辨中红外光谱观察到的蛋白质二级结构的变化。α-螺旋E和D插膜运动引起的别构效应驱动一对交叉排列的跨膜螺旋A和B发生剪切运动,导致附着于跨膜螺旋对上的叶绿素分子与相邻的叶黄素分子之间的间距随温度的升高或酸度的增加而变小。进一步通过量子化学计算,揭示叶黄素分子(LUT)暗态(S1态)和叶绿素a激发态电子态耦合强度在一定的温度范围内,也随温度升高而增强,导致叶绿素激发态的能量淬灭,并且能量淬灭主要发生在LUT-Chl612这一色素对上(图3)。
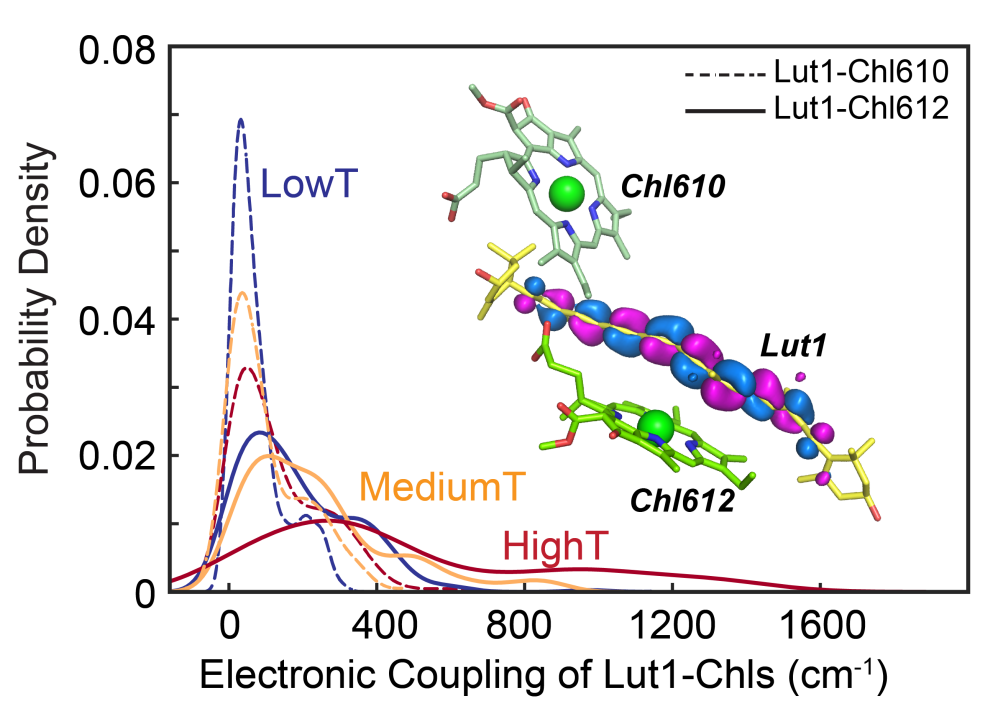
图3. 不同温度下LHCII三聚体中叶黄素分子1 (Lut1)和叶绿素分子间电子态耦合强度以及叶黄素分子的构像扭曲。
更为详细的图像是(图4),当310螺旋E转变为α-螺旋后,触发螺旋E和D在类囊体腔侧的插膜运动,并分别以螺旋D端的残基L206与螺旋B末端残基V80、L84构成剪叉内侧作用力的支点,以螺旋E端的残基W97与螺旋A在类囊体腔侧剪叉的末端残基F194、F195构成另一作用力的支点。类囊体腔侧的螺旋E与D在热能或酸度的驱动下形成一对顶杠,以螺旋A、B交叉点处的盐桥(Arg70-Glu180, Glu65-Arg185)为铰点,分别从跨膜螺旋A与B交叉面内侧将剪叉式排列的螺旋顶开,导致剪叉角度增加,带动两个剪叉式排列的叶黄素分子做相应的开叉运动,缩短了叶黄素分子与相邻的叶绿素分子之间的距离,增强了色素对间电子态的耦合,提升了叶绿素激发态能量的淬灭效率。尤其在低温条件下,位于腔侧的谷氨酸(E94)与螺旋E近端的赖氨酸K99形成氢键,随着温度的升高,螺旋E逐渐朝中心靠近,E94-K99之间的相互作用减弱,而E94与处于螺旋E中段的谷酰胺Q103形成新的氢键,这说明E94协同参与了螺旋E的插膜过程。上述模型与分子生物学中单点突变的实验结果相符合,如将E94突变成电中性甘氨酸,会导致叶绿素荧光淬灭效率降低。可见E94恰如开关中的触臂,K99及Q103分别为两个触点,当E94与K99相触的时候,系统处于高效捕光态,而当E94与Q103相触的时候,则处于光保护状态。
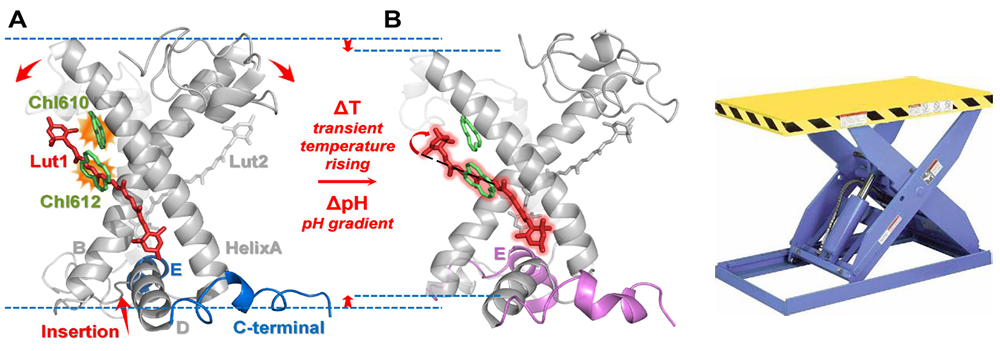
图4. 热能及酸度驱动LHCII三聚体分子开关剪叉式运动工作原理(左)及剪叉式升降机构(右)类比示意图。
可见,LHCII恰如剪叉式升降机一样的分子开关 (图4),在热能或酸度诱导的疏水作用下,触发螺旋E与D向疏水内核的插膜运动,将动力分别传给剪叉的两臂螺旋A与B,引起像剪叉式升降机一样的运动,迫使叶黄素分子和相邻的叶绿素分子相互接近,从而实现叶绿素激发态的能量淬灭。从分子机器的角度可得出以下三个推论:
(1)插膜运动速率是能量淬灭过程的速率决定步骤,时间尺度为1.24微秒,因此光合系统能量耗散也应该在这个时间尺度。文献报道的实验结果表明,活体叶片能量耗散的时间尺度为1.4微秒。
(2)从机械的角度考虑,腔侧螺旋插膜作用需要经过几道环节的传递才能够到转变为跨膜螺旋的剪叉升降运动,而传递过程中涉及的机械单元是典型的软物质,因而光合膜刚度越大,力的传递效率越高,能量耗散的效果也就越好,这点也为已知实验事实所证实。
(3)能量淬灭态对应的是交叉跨膜螺旋对顶角更为张开的状态,因而处于光保护状态的光合膜应当变薄(图4,左),这一点也与文献报道实验观测相符。
上述分子机制不仅与迄今报道的众多生物学实验观测相符,还揭示了植物是如何应用稳态温度和瞬态温度来实现高效能量耗散、同时又可以避免蛋白质热变性的。一般情况下,当环境温度为25℃时,叶片的温度可达40℃。叶片对激发能的耗散驰豫时间为10微秒量级,进一步估算10微秒内吸收的光子数全部变成热量可以引起LHCII三体瞬态温度上升14℃,并且该瞬态温度持续时间远小于100微秒的蛋白质开始变性的条件,因此不会引入额外的蛋白质变性。此时处于能量耗散状态的瞬态温度达到54℃,正好处于能量淬灭的最佳温度附近,而蛋白质的稳定性则由稳态温度40℃决定。可见自然界利用高瞬态温度实现最优化能量淬灭,同时维持较低的叶片稳态温度以保证蛋白质结构的完全可逆性。
该项工作以“Dynamical and Allosteric Regulation of Photoprotection in Light Harvesting Complex II”为题在线发表于Science China Chemistry. (https://doi.org/10.1007/s11426-020-9771-2)。中科院物理所博士生李昊和深圳湾实验室副研究员王英杰为论文的并列第一作者,翁羽翔和高加力为共同通讯联系人。
该研究得到了国家自然科学基金委重点项目(21433014)和中国科学院科研仪器设备研制项目(YZ200842,YJKYYQ20170046)的支持。分子动力学模拟工作分别得到了国家自然基金委、科技部重点研发计划高性能计算专项和美国NIH的资助。
原文链接:
https://link.springer.com/article/10.1007/s11426-020-9771-2
DOI:https://doi.org/10.1007/s11426-020-9771-2