

长期以来,光合捕光系统的传能机制都是由Förster 共振传能描述的,这种适用于给-受体色素分子间距离较远、相互作用较弱的经典传能模式不可避免的存在能量耗散。然而在实际捕光系统中,色素间的相互作用往往比较强,耦合比较大。这时色素之间会共享分子轨道,激发其中任何一个色素,它的激发态都会发生离域,形成这些色素分子激发态的叠加,即相干叠加态,也被称为激子态。这种相干叠加态的传能,称为相干传能。实验表明,相干态传能效率显著高于经典传能机制。实验中,受激系统大多是非孤立的,和环境热库作用导致位相关系的破坏,即发生退相干过程,而对于处于自然条件下的光合系统,所处的室温条件及介质分子(如蛋白质和水分子)又会加剧该退相干过程。对于电子激发态,退相干过程的时间尺度通常在几个到数十飞秒之间,远远小于光合作用色素间发生有效传能的数百飞秒时间范围。因此,如果光合系统存在有效的量子相干态传能机制,那么量子相干态的寿命必须和传能时间相匹配才能发挥作用。
2024年4月12日,Nature Communications 杂志在线报道了中国科学院物理研究所/北京凝聚态物理国家研究中心翁羽翔团队(SM6组)题为“激子-振动耦合能量耗散促成量子位相同步实现光合天线长寿命量子相干态”(Quantum Phase Synchronization via Exciton-Vibrational Energy Dissipation Sustains Long-lived Coherence in Photosynthetic Antennas)的研究工作。该团队与中国科学院烟台海岸带研究所秦松课题组合作,应用自行研制的二维电子光谱仪,研究了重组别藻蓝蛋白(Recombinant allophycocyanin,rAPC)的长寿命量子相干态及其量子位相同步机制。
别藻蓝蛋白(APC)是红藻和蓝藻中藻胆体复合物的核心捕光天线蛋白。APC能将从藻胆体杆部捕获的光能有效地传递至光系统的反应中心,其总体的量子效率超过 90%。图1展示了具有三重对称性APC三聚体的晶体结构。单体中色素α84和β84,距离约50埃。形成三聚体后,不同单体上的PCB色素距离约20埃,形成三对相同的α84-β84二聚体。该二聚体的电子耦合强度为155cm-1,形成的激子态能量劈裂约800cm-1,是研究二聚体激子态的理想光合天线样品。
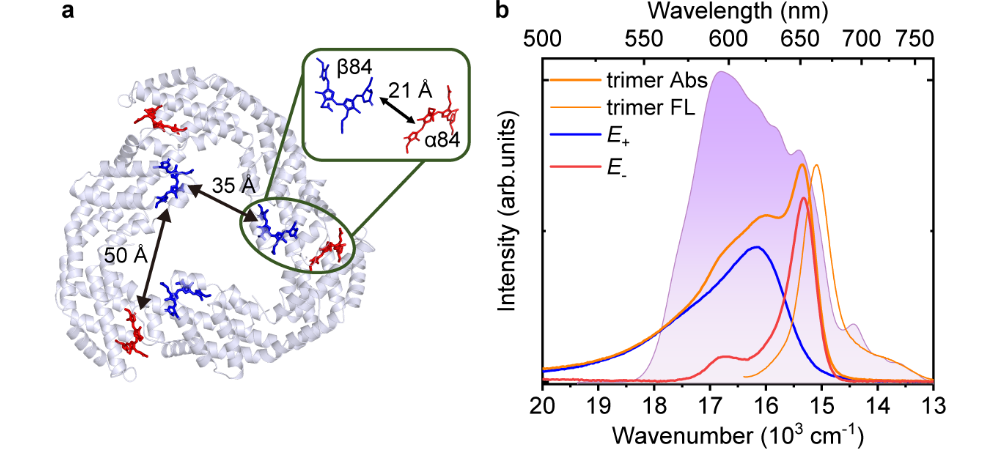
图1. APC的晶体结构和重组别藻蓝蛋白(r-APC)和光谱表征。(a)APC的晶体结构(PDB编号: 1ALL),其中相邻单体的α84(红色)、β84(蓝色); (b)r-APC三聚体的吸收(橙色粗线)和荧光光谱(橙色细线)以及激发脉冲的光谱范围(紫色阴影)。
二维电子光谱是一种同时具有高的时间分辨率和光谱分辨率的非线性光谱学方法,其基本原理是三光子回波测量。相干态激子导致不同激发路径之间的干涉即形成实验上可观测到的量子拍频现象,被认为是相干传能过程的确凿证据。
量子位相同步在经典物理学中的对应体是惠更斯双摆实验。受该实验的启发,国际上不同研究组建立了描述光合系统量子相干态的二聚体激子系统。理论研究表明,二聚体激子系统具有克服环境噪声涨落实现量子位相同步的内在机制。将二聚体中激子态-振动耦合系统的哈密顿量在离域化的激子态基矢下用 Jaynes-Cummings 形式重新表示,获得激子态-振动耦合表示:
其中ω为振动频率,θ为混合角,描述二聚体中激子离域化程度, σx为电子跃迁算符。可见,只有反对称的激发态和基态集体振动模能够与两个离域的激子态发生耦合。因此,当振动频率与两个激子态能级劈裂发生能量共振时,反对称集体振动模式的能量会被电子态的快速退相干过程( 30飞秒)耗散掉,只有对称集体振动模式能够被保留下来,由此导致了两个集体振动模式的位相同步(见图2)。
另一方面,在脉冲光激发下,分子的振动模式受布朗力作用进行欠阻尼振荡,并进一步导致激子能级的动态斯托克斯位移(ΔE_(t) )。通过引入集体振动坐标的初始相位φ,可以进一步得到低能激子态能级含时动态斯托克斯位移的解析表达式:
其中A0和γ为振动模的振幅和弛豫速率。上标i表示不同电子态。该公式表明激子能级的动态斯托克斯位移受集体振动模式的初始相位的调控。一旦集体振动模式变得关联,即φs-φa=0,上述公式表明该振动模对激子态下能级的动态斯托克斯位移贡献就会完全消失,也就是说该振动模不参与激子态下能级的能量耗散过程,从而也为激子-振动量子相干态提供了一种保护机制。
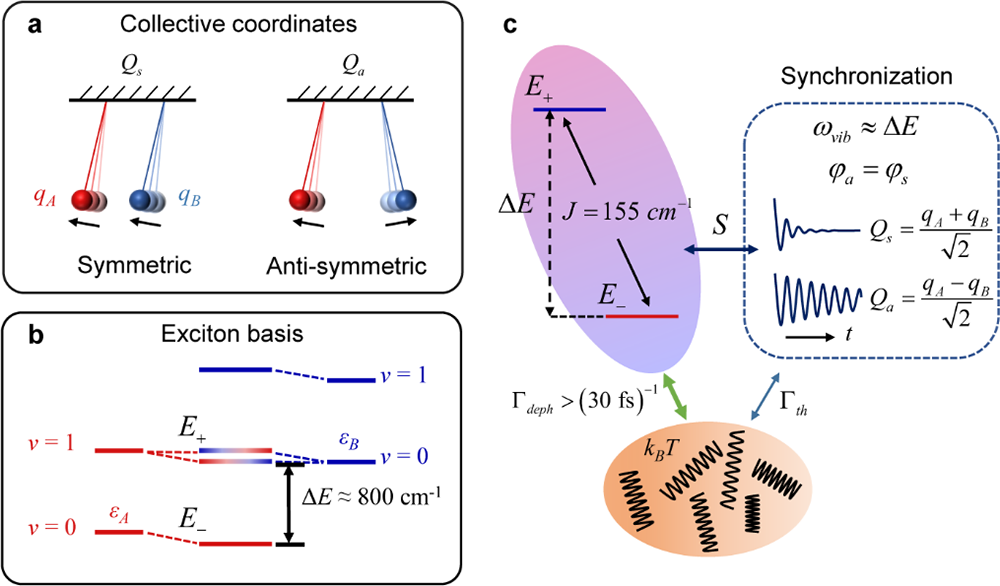
图 2. 量子位相同步机制的示意图。(a)利用一对钟摆的同相和反相运动示意二聚体中的对称和反对称集体振动模式。(b)电子-振动耦合的二聚体能级示意图。电子-振动混合的能级采用渐变色表示。(c)激子-振动-环境相互耦合的系统示意图。由于与激子态耦合,反对称集体振动模式的退相干寿命远短于对称集体振动模式。
基于上述二聚体激子-振动耦合理论,如果振动能量和激子能级劈裂能发生共振或者准共振时,二聚体的对称和反对称集体振动模就会实现位相同步,并可以获得以下实验可检验的推论:
(1)与单体色素分子相比,由于反对称集体振动模的耗散,与二聚体激子态能级劈裂发生共振的相干振动模强度只有单体中的一半。
(2)与二聚体激子态能级劈裂发生共振的振动模不参与低能激子态动态斯托克斯位移,即不参与能量耗散。
(3)与单体相比,二聚体电子-振动耦合量子相干态的寿命会延长。
上述三个理论预测分别为重组别藻蓝蛋白r-APC三聚体(含三对色素二聚体)和APC亚基(只含一个色素)的二维电子光谱所证实,如图3和4所示。
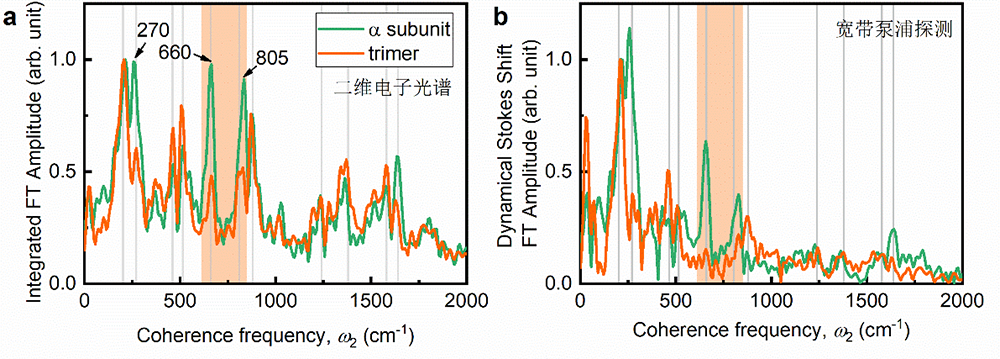
图3. 量子位相同步的实验验证。(a)由二维电子态相干光谱检测的积分相干谱揭示的r-APC三聚体在近共振频率区域(660和805cm-1)相干振动模振幅衰减一半效应。亚基(绿线);三聚体(橙线);(b)由宽带泵浦-探测光谱检测的动态斯托克斯位移相干谱,揭示660和805cm-1不参与激子最低能态的能量耗散。
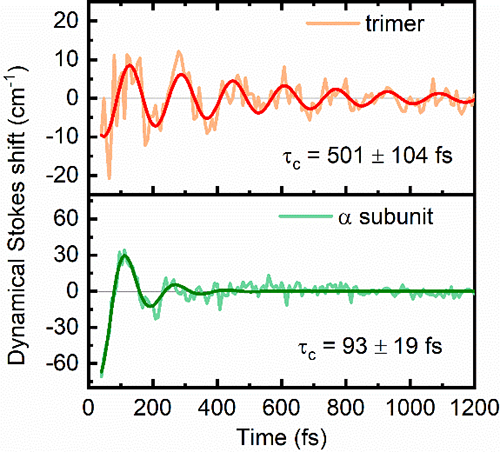
图4. 二维电子态相干光谱检测的三聚体和亚基的动态斯托克斯位移相干动力学衰减曲线。三体量子相干态寿命几乎是亚基的5倍,证实量子位相同步保护了电子-振动耦合量子相干态。
该工作从理论上阐明了量子位相同步的量子力学机制,预测了量子位相同步导致的实验现象,并通过二维电子态相干光谱实验定量地加以证实,从而揭示量子位相同步是二聚体激子通过电子-振动耦合,抵御环境噪声、保护长寿命电子-振动相干态的一种普适策略,是大自然应用量子力学优化传能路径的杰作。该原理不仅仅适用于光合体系,也必将为人工设计的量子相干系统所借鉴。
中国科学院物理所博士后朱锐丹,烟台海岸带所副研究员李文军,物理所博士后甄张赫为共同第一作者,烟台海岸带所秦松,物理所翁羽翔为通讯作者。该研究得到了国家自然科学基金委(T2350011, 22027802, U2230203和41906109),中国科学院(XDB33000000)和山东省自然科学基金委(ZR2021LLZ003)的支持。
原文链接:https://www.nature.com/articles/s41467-024-47560-6.pdf